


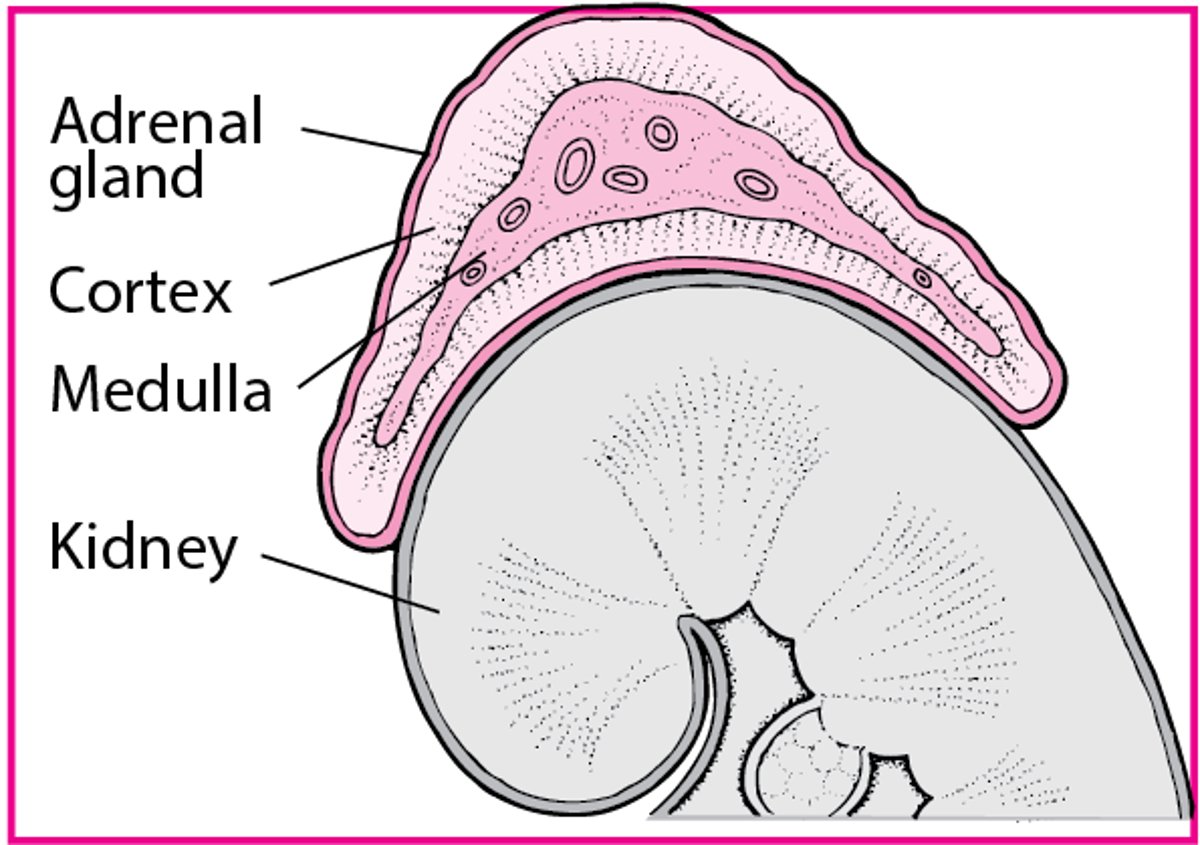
The adrenal glands, located on the cephalad portion of each kidney (see figure Adrenal Glands), consist of a
Cortex
Medulla
The adrenal cortex and adrenal medulla each have separate endocrine functions.
Adrenal Glands

Adrenal cortex
The adrenal cortex produces
Glucocorticoids (primarily cortisol)
Mineralocorticoids (primarily aldosterone)
Androgens (primarily dehydroepiandrosterone and androstenedione)
Glucocorticoids promote and inhibit gene transcription in many cells and organ systems. Prominent effects include anti-inflammatory actions and increased hepatic gluconeogenesis.
Mineralocorticoids regulate electrolyte transport across epithelial surfaces, particularly renal conservation of sodium in exchange for potassium.
Adrenal androgens’ chief physiologic activity occurs after conversion to testosterone and dihydrotestosterone.
The physiology of the hypothalamic-pituitary-adrenocortical system is further discussed elsewhere.
Adrenal medulla
The adrenal medulla is composed of chromaffin cells, which synthesize and secrete catecholamines (mainly epinephrine and lesser amounts of norepinephrine). Chromaffin cells also produce bioactive amines and peptides (eg, histamine, serotonin, chromogranins, neuropeptide hormones). Epinephrine and norepinephrine, the major effector amines of the sympathetic nervous system, are responsible for the response to stress (“flight or fight” response), which includes
Chronotropic and inotropic effects on the heart
Bronchodilation
Peripheral and splanchnic vasoconstriction with skeletal muscular vasodilation
Metabolic effects, including glycogenolysis, lipolysis, and renin release
Clinical syndromes
Most adrenal deficiency syndromes affect output of all adrenocortical hormones. Hypofunction may be primary (malfunction of the adrenal gland itself, as in Addison disease) or secondary (due to lack of adrenal stimulation by the pituitary or hypothalamus). Some experts refer to hypothalamic malfunction as tertiary.
Adrenal hyperfunction causes distinct clinical syndromes depending on the hormone involved:
Hypersecretion of glucocorticoids results in Cushing syndrome.
Hypersecretion of aldosterone results in hyperaldosteronism.
Hypersecretion of androgens results in adrenal virilism.
Hypersecretion of epinephrine and norepinephrine results in symptoms of pheochromocytoma.
These syndromes frequently have overlapping features.
Hyperfunction may be compensatory, as in congenital adrenal hyperplasia, or due to acquired hyperplasia, adenomas, or adenocarcinomas.






